
Mouth breathing in Tursiops truncatus:
sound, video and audio recording.
Roberto Rutigliano, Sabrina Capelletti, Micaela Bacchetta,
Centro Ricerca Cetacei, Portoferraio Li, Italy.
Abstract
We will describe rare respiration behavior in an adult Bottlenose Dolphin (Tursiops truncatus) observed in 2011 near South coasts of Elba Island (Mediterranean Sea, Italy), which appears to breath via its mouth (fig.1A e fig.1B) rather than using its blowhole. This is the second case of such behavior in Cetacea described in literature supported by photo/video documentation, after Dawson’s published work et al. (2017) about an adult of New Zealand dolphin (Cephalorhynchus hectori) breathing mostly via its mouth. The decision to postpone the sharing of this important observation with the scientific community arises from the desire to enrich our work with other data on new sightings of this dolphin and its behavioral flexibility to deal with a probable pathological problem.
Detailed observations were made on 28 June 2011 and on 1 July 2011 and consisted in 10 photographs surfacing sequences and a 49-seconds’ video with sound evidence of the inspiration noise clearly audible as the dolphin opened its mouth.
The dolphin was observed in two groups around the South coasts of Elba Island about 3 miles off Calamita Mount (fig. 1C).
We want to present the results of our observations concerning such uncommon events, trying to explain them and give a general overview of the breathing via mouth in Cetacea from an anatomic point of view, starting from the theoretical assumption that the anatomy of Cetacea is based on the analysis and dissection of cadavers, mainly Mysticeti.
 Figure 1. Surfacing breaths in dolphin tTS167 (A, B). Sightings map (C).
Figure 1. Surfacing breaths in dolphin tTS167 (A, B). Sightings map (C).
Introduction
The evolutionary path and the adaptations to the aquatic environment have led the Cetacea to substantial changes in their morphology and physiology, keeping distinctive features of terrestrial Mammalia: first of all, the breathing of atmospheric air for oxygenation of their body.
Therefore, the airways had to change and move in order to allow horizontal and sinusoidal swimming even during breathing. The nostrils, in fact, have moved to the apex of the skull to form the blowhole, which in Odontoceti is fused in a single nostril. The opening of the breather occurs by voluntary muscles, as well as the expansion of the thoracic cavity, so, unlike terrestrial mammals, all Cetacea decide when to breathe.
 Figure 2. Normal inspiration by another member of the same group.
Figure 2. Normal inspiration by another member of the same group.
Furthermore, the upper airways had to change their anatomical position to follow the migration of the nostrils, so in the Bottlenose Dolphin, as in most Cetacea, the larynx, with an elongated beaked epiglottis, passes through the esophagus pointing upwards towards the blowhole separated by the nasopharynx and by the air sac system. Simultaneously, the soft palate extended to the larynx, in order to seal and separate the oropharynx from the nasopharynx, preventing the passage of food or water from the mouth to the airways.
The trachea is proportionally short as well, nevertheless wider than normal, due to the reduced length of the neck itself. Thus, it was possible to reduce space and ensure the emission of large volumes of air at a higher speed during expiration.
Cetaceans therefore appear to have a completely separate digestive system and respiratory system, breathing and feeding at the same time.
However, it seems that the larynx can voluntarily withdraw from the palato-pharyngeal sphincter if necessary (Dawson et al., 2017). It has been observed in several captive individuals of different species of Odontoceta that the larynx had displaced to the left side during veterinary maneuvers to pull out non-food material or during intubation by artificial ventilation. The larynx displacement took place with initial resistance by the respiratory muscles and then returned to the normal intranarial position once the mechanical action ceased (Ridgway 1965; Dawson et al., 2017).
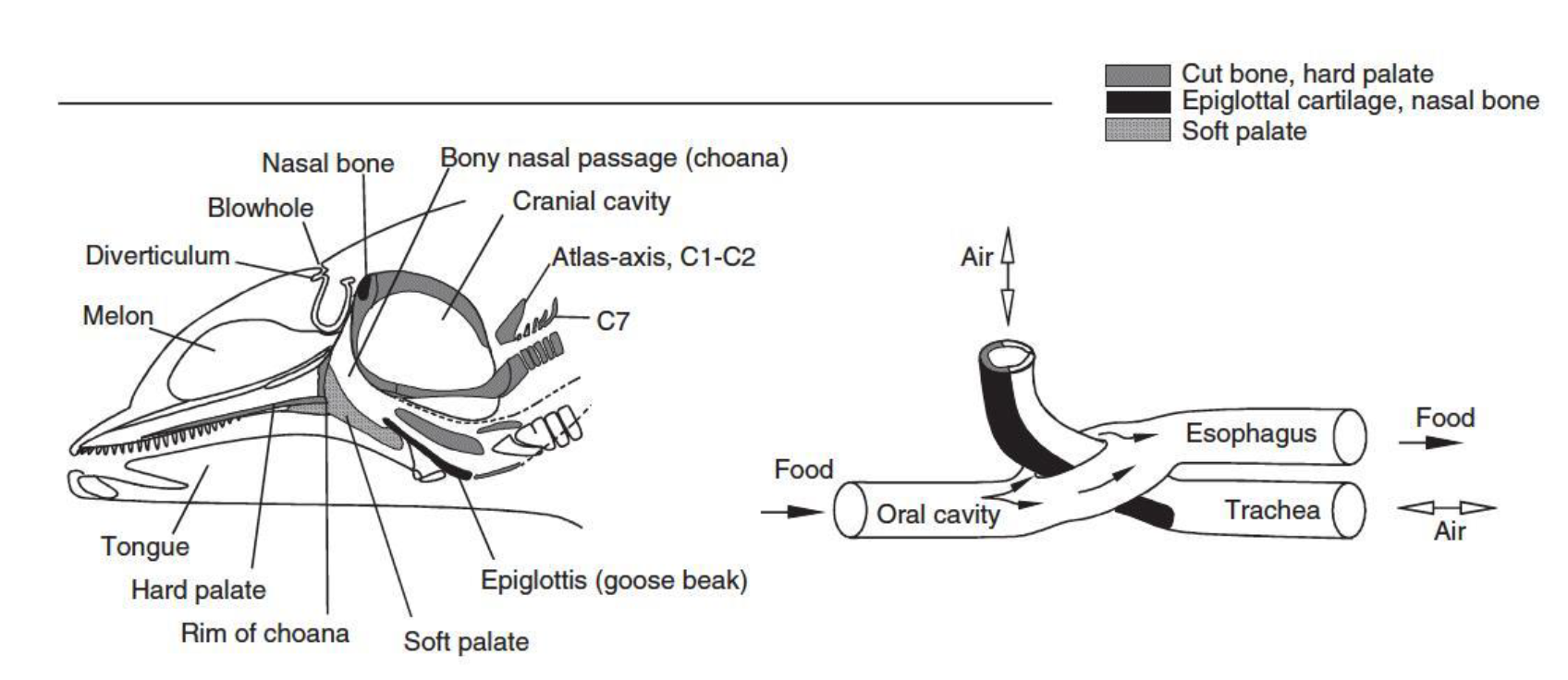
Figure 3. Anatomical detail in Odontoceta (Source: Encyclopedia of marine mammals, 2009)
Rarely, at the end of the unblocking or intubation surgery of an individual, the respiratory muscles (intercostal, diaphragm, rectus abdominis and sternohyoid muscles; Cotten et al., 2008) were observed to respond for a few seconds faster than the nasal muscles: the individual opens its mouth first and inhales air here one/three times withdrawing the larynx from the palato-pharyngeal sphincter, until normal breathing is resumed through the blowhole.
It therefore appears that the larynx is not located in a fixed and permanent position but can be voluntarily dislocated by the animal itself when it ingests a large prey or manually by a doctor when intubating is required (Dawson et al., 2017).
However, several cases of fatal suffocation in Delphinidae (particularly in Bottlenose dolphins, Stolen et al., 2013), due to the ingestion of large prey, suggest that the dislocation of the larynx may have structural and spatial limits.
Materials and methods
Daily research activity and photo-ID surveys were conducted on sailing/motor boats from May to October in the whole Tuscan Archipelago, especially around Elba Island.
The photos and the video were obtained using a Nikon D300 camera, 18-70mm nikkor 1: 3.5 G and 70-200mm Af-s Vr nikkor 1: 2.8 Ge lenses; JVC hybrid camera Gc-px 10e 36mbps.
We use the photo-identification technique to recognize and follow animals during the years, with a matching simplification tool included in our online research data management software. This database is on the web and allows us to get reports about animal behavior with any computer means everywhere.
The individual dolphin studied in this article, never observed in previous sightings, was photo-identified and registered by the name "tTS167" (Northern Tyrrhenian Bottlenose Dolphin n.167).
The first observation of tTS167 on 28 June 2011 started at 11:41 am at the point of coordinates 42°40'23.196"N, 10°20'27.384"E (Lat.42.67311, Long.10.34094 in decimal degrees WGS84) and ended at 13:00 in the point of coordinates 42°41'21.552"N, 10°24'28.109"E (Lat. 42.68932, Long.10.407808 in decimal degrees WGS84). Water depth was 103 m and sea surface temperature 24°C. The individual tTS167 was observed in a pod of nine, with adults, sub-adults and a calf, in a compact circle formation.
The second sighting (without video material) on 01.07.2011 started at 09:50 at the point of coordinates 42°40'37.848"N, 10°19'39.576"E (Lat.42.67718, Long.10.32766 in decimal degrees WGS84) and ended at 10:40 in the point of coordinates 42°40'37.092"N, 10°19'38.928"E (Lat.42.67697, Long.10.32748 in decimal degrees WGS84). Water was 88 m deep and sea surface temperature was 24°C. The individual tTS167 was observed in a pod of seventeen, with adults, sub-adults and two calves, in a dispersed formation.
In the next ten years, during photo ID surveys particular attention was paid to meet the individual tTS167 again, in order to collect new data about the damage to its respiratory system.
The choice of the decade as a useful period was made by calculating a rational period in which a significant number of non-resident dolphins tend to be sighted at least a second time. In practice we have considered both the distances between 2 sightings of the same individual on all the individuals in the archive, and the same data relating only to individuals considered rare: i.e. with less than 6 total sightings, but more than 1, having the first sighting dated no later than 2011 and the second at least in the following calendar year.
Results
Photo-ID pictures were taken in both sightings, while videos were recorded only during the first in a complex situation without interaction of the group with the boat, especially for dolphin tTS167. The images and the videos show this dolphin breathing using its mouth and not its blowhole during surfacing, at an irregular breathing rate (wide and variable times between 2 puffs), which agrees on the existence of a pathology.
As to all other physiological and behavioral aspects, the dolphin appeared normal and in good physical condition. No evidence of external lesions on the blowhole or in the nasal region were detected from the frames analysis.
During direct observations the sound of a deep breathing was clearly heard as the dolphin opened its mouth and no typical puff was visible from its blowhole. This was also confirmed in video analysis.
In every surfacing observed, tTS167 dolphin approached the surface at a steep angle: no classic arc sequence (melon-blowhole-vaporized breath-dorsal fin) but the head emerged higher than normal, at a 30° angle from the water surface and always opening its mouth.

Figure 4. Breathing sequence in dolphin tTS167
Furthermore, in the first breath in the video, and less in the others, a puff of water is quite evident in front of the dolphin's mouth, revealing pressurized air exhaled from the mouth. This would confirm the ability of the Cetaceans to create a physical bypass of its laryngeal closure using the palatopharyngeal sphincter.
In both sightings the two groups were observed in opportunistic feeding activities following a trawler. The ”mouth-breathing” dolphin showed breaching and competitive type interactions with other individuals who joined the hunt. Individual tTS167 reacted with a long series of jumps, always keeping oral breathing, as photo and video evidence show.
The individual was determined to be female due to the ethological habits of the Bottlenose dolphins. These dolphins usually form big groups of females and calves and few units of males, although it has been observed that the males can join females groups during opportunistic hunting.
The estimated age of tTS167 is about 10 years due to its body size and low amount of scratches and notches on its body and dorsal fin, featuring a young but mature animal.

Figure 5. Other pictures about breath via mouth in dolphin tTS167
We chose to wait ten years for a new sighting of tTS167 because for the 185 identified Bottlenose dolphins sighted for the first time until 2011 only 2 were re-sighted more than ten years later (i.e. 11 and 12 years). Only once a dolphin was seen 10 years later; 2 after 9 years; 2 after 8 years, with increasing effect at a decreasing time distance.
Furthermore, taking a sample of 22 rare individuals (using the criteria described above) their average of the meeting distance is 5.6 years.
So, the choice of 10 years seemed a good period respecting the scientific importance of the event described.
Unfortunately, the individual tTS167 has not been seen in this time lapse.
Database analysis showed that in the first sighting 4 other dolphins were photo-identified and in the second some 3 more. Only one dolphin was present in both sightings, named tTS166. All of these individuals have been seen and recognized in the future at least once after one or more years. Dolphin tTS166 has been seen again nine more times during the next 3 months and never again.
Conclusions
Since it is a free individual and we cannot ascertain the causes of its unusual breathing, we hypothesize that in tTS167 the passage of air from the mouth is therefore determined by modifications in the anatomy of its respiratory system, due to various causes limiting the flow of air through its blowhole:
- A congenital dysfunction of the muscles that open the blowhole.
- An occlusion somewhere in the upper respiratory tract due to a foreign body or trauma.
- A clinical lesion with cyst or secretion occlusion following a viral, bacterial or parasitic infection.
In the literature there is a single documented case of oral respiration in cetaceans, in particular in Delphinidae (Dawson et al., 2017), that suggests similar causes. Furthermore, there are several sightings and testimonies reported online, even in other species, which support the casuistry.
It seems impossible to give greater responsibility to any of the listed causes, by virtue of the fact that the activity and good general condition of tTS167 during both sightings indicate that the respiratory system ensured effective and efficient breathing for survival, at least in the short term.
The irregular respiratory rate and poor sociability with the boat are due more to a consequence of its disease than to any cause.
In fact, the quality of the air entering orally does not allow optimal oxygenation of the blood, moreover, having no more air in the natural upper respiratory tract close to the melon, such as the air sacs, the important communicative vocalizations and those suitable for biosonar are reduced, causing the dolphin to lose control of the three-dimensional space around it.
In the long term, this could become the biggest handicap of a toothed dolphin that uses the mouth to convey the air, compromising its nutrition and its sociability, which could be its greatest resource, due to the ability of the dolphins to use the bouncy sounds from the vocalizations of the companions to move in space (collective vision).
In recent years, we have been waiting for the individual to meet again in order to verify its health conditions after some time. Unfortunately, this did not happen and, although this would suggest the worst in relation to the observed event, the strong possibility that tTS167 is a transient or untracked dolphin is feasible, also considering that the other 22 individuals in the database, out of the 396 total in 2021, which have been sighted and photo-identified only once, also due to fins with little characterizing markers, therefore susceptible to changes and recognition errors.

Figure 6. Right and left side of dorsal fin in tTS167
The fact that all the other photo-identified Bottlenose dolphins in the two sightings have been seen at least one other time does not support the hypothesis that the individual may have survived the disease, on the other hand it does not exclude it.
We at the Centro Ricerca Cetacei can only keep on committing ourselves to try to identify tTS167 again in the event of a new meeting and communicating it promptly to the scientific community.
Literature cited
Cotten P.B., Piscitelli M.A., McLellan W.A., Rommel S.A., Dearolf J.L., Pabst D.A. (2008). The gross morphology and histochemistry of respiratory muscles in bottlenose dolphins, Tursiops truncatus. J. Morphol., 269: 1520–1538. doi:10.1002/jmor.10668
Dawson S.M., Fordyce R.E., Ridgway S.H., Brough T.E., Slooten E. (2017). Observations of a New Zealand dolphin (Cephalorhynchus hectori) breathing via its mouth. Mar Mam Sci, 33: 350–355. doi:10.1111/mms.12349
Ridgway S.H., (1986). Diving by cetaceans. Diving in animals and man. The Royal Norwegian Society of Science and Letters, Trondheim, Norway, pp.33-62.
Stolen M., St. Leger J., Durden W.N., Mazza T., Nilson E. (2013). Fatal Asphyxiation in Bottlenose Dolphins (Tursiops truncatus) from the Indian River Lagoon. PLoS ONE 8(6): e66828. https://doi.org/10.1371/journal.pone.0066828
Würsig B., Perrin W.F., Thewissen J.G.M. (2009). Encyclopedia of marine mammals. 2° edition. ISBN 0080919936, 9780080919935. Academic Press.